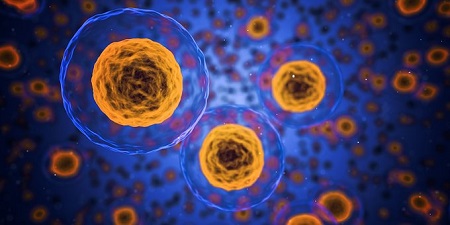
La reprogramación celular es la tecnología que nos permite obtener células madre pluripotentes inducidas o iPSC (del inglés, induced pluripotent stem cells). Al tener todas las ventajas y capacidades de las células madre embrionarias y eludir sus problemas éticos y biológicos, el descubrimiento y desarrollo de las iPSC se ha convertido en uno de los avances más importantes en el campo de la biología en las últimas dos décadas. El proceso consiste en la expresión de los denominados factores de Yamanaka, Oct4, Sox2, Klf4 y c-Myc (OSKM); cuatro factores de transcripción que permiten la reconversión del destino celular somático a un estado pluripotente similar al de las células madre embrionarias. Dadas las importantes diferencias entre ambos estados celulares, esta transformación supone una profunda reorganización del fenotipo celular a todos los niveles. Para ello, las células somáticas deben sortear una serie de barreras celulares y moleculares que impiden la reconversión celular. Durante este proceso, las células han de ser capaces de estabilizar la pluripotencia mediante la reactivación de su programa de control endógeno.
En el grupo del profesor Josema Torres de la Unidad de Neurobiología Celular en la Universidad de Valencia, llevamos años estudiando los mecanismos celulares y moleculares que controlan la fase plástica de la reprogramación celular. Desde el inicio de este reciente campo de investigación, múltiples nuevas combinaciones de diferentes factores de transcripción y compuestos han demostrado ser capaces de permitir la reprogramación celular. Concretamente, se ha descrito que la expresión exógena de c-Myc no es necesaria, aunque sí beneficiosa, para la reprogramación celular (Knoepfler, 2008). Estos hallazgos han reforzado la idea de que esta proteína no desempeña ningún papel crucial en el proceso de reprogramación celular. Sin embargo, el pasado noviembre nuestro grupo publicó un estudio en Stem Cell Reports en el que describimos algunas de las funciones indispensables que realiza la proteína c-Myc a este nivel (Prieto et al. 2018), demostrando la necesidad, al menos de manera endógena, de esta proteína para obtener iPSC. Concretamente, c-Myc dirige la reorganización mitocondrial e induce un programa energético híbrido para favorecer la transición fenotípica a la pluripotencia.
Las células somáticas adquieren un estado plástico conforme superan las barreras celulares de la primera fase del proceso. Este fase se caracteriza, entre otras cosas, por experimentar una notable remodelación de su morfología mitocondrial y una reconversión de su metabolismo celular (Folmes et al., 2010; Prieto et al., 2016). Se ha sugerido que esta transición metabólica es un fenómeno sincrónico en el que un aumento gradual en el flujo glucolítico ocurre en paralelo a una reducción de la respiración celular durante el proceso de reprogramación (Wu et al., 2016). Sin embargo, en este trabajo describimos que c-Myc activa de manera temprana un fenotipo metabólico híbrido, marcando las células propensas a la reprogramación celular.

Figura 1. Las células somáticas adquieren un estado plástico durante la reprogramación celular caracterizado por una morfología mitocondrial fragmentada, un metabolismo híbrido y un alto potencial de membrana mitocondrial.
La regulación al alza de las enzimas glucolíticas y de la fosforilación oxidativa (OXPHOS, del inglés, oxidative phoshorylation) por c-Myc puede explicar el aumento temprano y en paralelo de ambas rutas metabólicas observado durante la reprogramación celular. La expresión de OSKM causa un pico de activación temprana y transitoria de OXPHOS (Kida et al., 2015) que está inducido por la actividad combinada de OSK y c-Myc. La inhibición pronunciada de OXPHOS tras este pico de activación a niveles casi somáticos coincidió con la regulación al alza de los flujos glucolíticos. La dinámica opuesta de estas dos vías podría sugerir una regulación negativa de OXPHOS por altas tasas metabólicas glucolíticas, lo que limitaría la disponibilidad de piruvato en las mitocondrias para maximizar el reciclaje de NAD+. Sin embargo, no se pudo descartar una regulación orquestada de ambas vías por la interferencia de diferentes programas de expresión génica inducidos por los factores de reprogramación. En este sentido, descubrimos una regulación positiva de la proteína Atpif1 por c-Myc, la cual puede jugar un papel en la modulación de la actividad de OXPHOS.
Aunque es necesario para la reprogramación celular (Kida et al., 2015; Prieto et al., 2018), se desconoce si este pico de OXPHOS es únicamente un subproducto de la expresión forzada de los factores de reprogramación o si desempeña un papel específico en el proceso de plasticidad celular. En este sentido, es posible que el aumento de OXPHOS pueda inducir una explosión de especies reactivas de oxígeno (ROS, del inglés, reactive oxygen species) durante la reprogramación celular. Como el ROS puede activar vías de señalización pro-proliferación (Hawkins et al., 2016), un aumento temprano y transitorio en los niveles de ROS podría estimular la proliferación celular y, por lo tanto, ser favorable para la reprogramación celular. En este sentido, se ha encontrado que existe una señal de ROS óptima para la reprogramación celular (Zhou et al., 2016). Sorprendentemente, nuestras observaciones muestran que las células que muestran un alto potencial de membrana mitocondrial (MMP, del inglés, mitochondrial membrane potential) son propensas a la reprogramación celular, revelando el importante papel de la función mitocondrial en este proceso. Estas células con un alto MMP mostraron tanto un aumento de la glucólisis como una alta tasa de OXPHOS, lo que indica que se puede establecer fácilmente un metabolismo híbrido durante las primeras etapas de la reprogramación celular, probablemente debido a la acción de c-Myc.
Este trabajo, junto a un estudio previo de nuestro grupo (Prieto et al., 2016), refuerza la idea de la existencia de importantes similitudes entre la reprogramación celular y la tumorigénesis celular. Existen indicios que sugieren que la función metabólica de las mitocondrias, su dinámica y el ROS juegan papeles similares, indicando que los intermediarios celulares de ambos procesos pueden presentar una dinámica plástica parecida (Apostolou and Hochedlinger, 2013; Prieto and Torres, 2017).
Referencia:
Prieto J et al.. MYC Induces a Hybrid Energetics Program Early in Cell Reprogramming. Stem Cell Rep. 2018, 11, 1-14. Doi: https://doi.org/10.1016/j.stemcr.2018.10.018
Bibliografía:
Apostolou E and Hochedlinger K (2013). Chromatin dynamics during cellular reprogramming. Nature. 2013, 502, 462-471. Doi: https://doi.org/10.1038/nature12749
Folmes CD et al. Somatic oxidative bioenergetics transitions into pluripotency-dependent glycolysis to facilitate nuclear reprogramming. Cell Metab. 2011 Aug 3;14(2):264-71. doi: 10.1016/j.cmet.2011.06.011.
Hawkins KE et al. NRF2 orchestrates the metabolic shift during induced pluripotent stem cell reprogramming. Cell Rep. 2016 Mar 1;14(8):1883-91. doi: 10.1016/j.celrep.2016.02.003.
Kida YS et al. (2015). ERRs mediate a metabolic switch required for somatic cell reprogram- ming to pluripotency. Cell Stem Cell 2015 May 7;16(5):547-55. doi: 10.1016/j.stem.2015.03.001.
Knoepfler PS. Why myc? An unexpected ingredient in the stem cell cocktail. Cell Stem Cell. 2008 Jan 10;2(1):18-21. doi: 10.1016/j.stem.2007.12.004.
Prieto J, et al. Early ERK1/2 activation promotes DRP1-dependent mitochondrial fission necessary for cell reprogramming. Nat Commun. 2016 Mar 31;7:11124. doi: 10.1038/ncomms11124.
Prieto J, Torres J. Mitochondrial Dynamics: In Cell Reprogramming as It Is in Cancer. Stem Cells Int. 2017;2017:8073721. doi: 10.1155/2017/8073721.
Wu J, et al. Cellular Metabolism and Induced Pluripotency. Cell. 2016 Sep 8;166(6):1371-1385. doi: 10.1016/j.cell.2016.08.008.
Zhou G, et al. Optimal ROS Signaling Is Critical for Nuclear Reprogramming. Cell Rep. 2016 May 3;15(5):919-925. doi: 10.1016/j.celrep.2016.03.084.
Javier Prieto1, Marian León2 y Josema Torres2,3
1 Gene Expression Laboratory, Salk Institute for Biological Studies, 10010 North Torrey Pines Road, La Jolla, CA 92037, USA.
2 Dpto. Biología Celular, Biología Funcional y Antropología Física, Universidad de Valencia, 46100 Burjassot, España.
3 Instituto de Investigación Sanitaria INCLIVA, 46010 Valencia, España.
El gen AtCDF3 promueve una mayor producción de az...
Un estudio con datos de los últimos 35 años, ind...
Un equipo de investigadores de la Universidad Juli...
En nuestro post hablamos sobre este interesante tipo de célula del si...
Palobiofarma S.L. se complace en anunciar la finalización del tratami...










