Did you know that living primates include between 200 and 450 species (depending on the classification system used) and primarily live in the tropical latitudes of Africa, Asia, and the Americas? Primates includes humans and all non-human primate (NHP) species.
Why do we use NHPs in reproductive research?
Due to their phylogenetic closeness to humans (Fig. 1) with regards to anatomy, endocrinology and reproductive physiology, NHPs are invaluable research models for studies on fertility/infertility, IVF or ART procedures, male contraception and reproductive toxicology. Several NHP species have been highlighted as key animal models in human-related studies and are widely used, including Old Word monkeys (macaques, vervet monkeys and baboons) and New World monkeys (squirrel and owl monkeys). Although hominoid species (great apes) are probably closest to humans, the use of these animals in biomedical research has been opposed worldwide.

Fig. 1: Classification of primates indicating their phylogenetic closeness to humans.
Species comparisons and standardization
When using NHPs in studies to relate the results back to what would possibly happen in humans, one should be careful not to compare apples to oranges. In order to avoid this, two pitfalls must be highlighted. Firstly, just because certain species are phylogenetically close, it cannot be assumed that their semen and sperm characteristics are similar (though it often is!). Secondly, species comparisons require objective, consistent and sensitive techniques to identify differences among species, especially in treatment studies to recognize compromised sperm functions. In both of the aforementioned instances, CASA can be an extremely helpful tool to generate comparable baseline data, on condition that standardised protocols and media are used in preparation of sperm samples and species-specific cut-off values are employed in the CASA settings.
Using SCA in diverse NHP studies
A. Sperm structure and function
When we compared sperm structure and function of seven mammalian species, sperm morphometry and motility characteristics (mainly obtained with SCA software) were closely related among three NHP species (Fig. 2 & 3). Although human sperm share some structural aspects with the NHPs (Fig. 2), its swimming characteristics were closer to that of merino rams and mice than to the NHPs (Fig. 3).

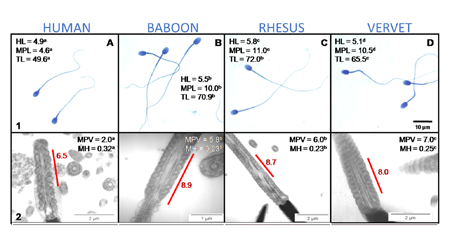
Fig. 2: Comparison of sperm morphometry parameters among human and NHP sperm. HL, = head length; MPL, midpiece length; TL, tail length, MPV, midpiece volume; MH, mitochondrial height.

Fig. 3: Cluster analysis of eight sperm motility parameters captured at 50 frames/second with SCA illustrating the clustering of species on the vertical axis and indicating the relatedness among the mammalian species on the horizontal axis (linkage distance). HS, Homo sapiens (human); PU, Papio ursinus (chacma baboon); RM, Macaca mulatta (rhesus monkey); CA, Chlorocebus aethiops (vervet monkey); OO, Ovis orientalis (merino ram); MM, Mus musculus (house mouse).
B. Comparing physiological media
Different physiological media used for sperm preparations seem to affect sperm performance in ART. While media to be used for human sperm assessment are generally prescribed in the WHO’s laboratory manual, the selection of an appropriate medium for NHP sperm functional testing are less clear. CASA can be useful in determining the effect of media on both percentage total sperm motility as well as several sperm subpopulations (Fig. 4).

Fig. 4: Comparison of five physiological media on sperm motility percentages in vervet monkeys. Red, human tubal fluid (HTF); blue, capacitating HTF; white, Ham’s F10; green, HD Sperm Wash; black, HD Sperm Capacitation (From: Mabotha, L (2019). Evaluation of sperm functionality in non-human primates, focussing on sperm capacitation. MSc Thesis, University of the Western Cape).
C. Toxicological studies
For any animal model to be relevant for reproductive toxicity testing, the test extract must produce a similar in vitro pharmacokinetic effect as found in humans. We found a similar effect on various sperm parameters when exposing human and NHP sperm to two heavy metals acting as environmental contaminants (CuSO4 and CdCl2). Not only was sperm swimming speed (VCL) affected by increased concentration of CuSO4 (Fig. 5), it also effected the induction of sperm hyperactivation (as measured by applying species-specific SCA cut-off values, Fig. 6).

Fig. 5: Effect of various concentrations of CuSO4 on curvilinear velocity of vervet monkey sperm as analysed by SCA at 50 frames/second.

Fig. 6: Hyperactivation induced in vervet monkey sperm after exposure to 5 mM caffeine and 100 µg/ml. (from Prag, F (2017). Evaluation of standard and development of new sperm functional tests in selected primate species. MSc Thesis, University of the Western Cape).
In conclusion, it is clear that CASA should play an essential part in any human-related reproductive studies where NHPs are used as animal models. CASA provides numerous applications for assessment of NHP sperm that can assist with both the standardisation of protocols/techniques and investigating the effect of external factors on sperm characteristics.
Dr Liana Maree (PhD)
Comparative Spermatology Laboratory
University of the Western Cape

La mejor actitud que podemos adoptar es la de trat...

The research team observed changes in head circumf...
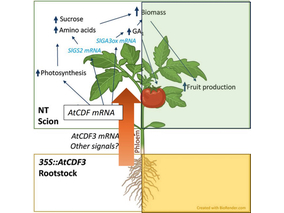
AtCDF3 gene induced greater production of sugars a...
En nuestro post hablamos sobre este interesante tipo de célula del...

Telum Therapeutics, a biotechnology company leveraging proprietary met...