Extracellular vesicles (EVs) transport proteins, lipids and nucleic acids. Therefore, they are of great interest as sources of biomarkers and as mediators of intercellular communication. However, when analyses are performed in serum or plasma, we face a problem that has been underestimated for years: lipoproteins.
This is not a trivial issue. Lipoproteins and EVs share physical properties such as size, density, and lipid composition, which means they co-isolate with most standard methods. The consequence? Many results attributed to EVs may be partially, or even predominantly, influenced by lipoproteins, compromising the biological interpretation of the data (1,2).
Recognising and addressing this problem is not a minor technical detail; it is an essential requirement for obtaining reliable and reproducible results.
Several types of lipoproteins circulate in blood, with sizes ranging from ~5 nm to more than 1,000 nm. This range overlaps extensively with that of EVs, typically described as 30 to 1,000 nm (3,4).
The key difference, and this is where the real problem begins, is quantitative. Lipoproteins are several orders of magnitude more abundant than EVs in plasma. It is estimated that the number of lipoprotein particles can exceed that of EVs by 20- to 100-fold, particularly in the case of HDL (4). In this context, particle-counting techniques simply detect “what is most abundant”.
Several studies have shown that widely used methods such as differential ultracentrifugation, density gradients or size-exclusion chromatography (SEC) do not efficiently separate EVs from lipoproteins (3, 5). It has even been demonstrated that co-purified LDL particles display sizes and behaviours that are virtually indistinguishable from EVs, and can react with antibodies commonly used as vesicle markers (1). This phenomenon creates a false sense of specificity: what is interpreted as an EV signal may correspond to lipoproteins masquerading as vesicles.
Lipoproteins have a direct impact on analytical techniques:
1. Particle counting and size distribution
Techniques such as Nanoparticle Tracking Analysis (NTA) or TRPS do not distinguish between EVs and lipoproteins. As a result, particle counts are often inflated and size distributions distorted, especially in HDL-rich samples (5).
2. Proteomic analysis
The presence of apolipoproteins, particularly ApoA1 and ApoB, significantly alters proteomic profiles obtained from EV preparations, masking low-abundance proteins that are genuinely associated with vesicles (4).
3. RNA and miRNA studies
Lipoproteins also carry RNA, including miRNAs. Comparative studies have shown that different isolation methods generate markedly different miRNA profiles, largely due to the co-purification of lipoproteins (6). This introduces a critical bias into biomarker studies.
In clinical studies, this issue becomes particularly relevant. Observed changes in EV “load” or molecular content may reflect variations in lipid metabolism rather than pathological processes related to EVs themselves (3, 5). Without proper control of lipoproteins, the risk of assigning biological significance to technical artefacts is substantial.
Strategies to mitigate the problem
No solution is perfect, but ignoring the problem is no longer an option. Proper separation is not a luxury. It is the difference between measuring signal… or simply well-dressed noise.
1.Sódar, B. W., et al. (2016). Low-density lipoprotein mimics blood plasma-derived exosomes and microvesicles during isolation and detection. Scientific Reports, 6, 24316.
2.Simonsen, J. B. (2017). What are we looking at? Extracellular vesicles, lipoproteins, or both? Circulation Research, 121(8), 920–922.
3.Yuana, Y., et al. (2014). Co-isolation of extracellular vesicles and high-density lipoproteins using density gradient ultracentrifugation. Journal of Extracellular Vesicles, 3, 23262.
4.Karimi, N., et al. (2018). Detailed analysis of the plasma extracellular vesicle proteome after separation from lipoproteins. Cellular and Molecular Life Sciences, 75, 2873–2886.
5.Mørk, M., et al. (2016). Particle size analysis of extracellular vesicles and lipoproteins in plasma using nanoparticle tracking analysis. Journal of Extracellular Vesicles, 5, 31638.
6.Buschmann, D., et al (2018). Evaluation of serum extracellular vesicle isolation methods for profiling miRNAs by next-generation sequencing. Journal of Extracellular Vesicles, 7(1), 1481321.
7.Lobb, R. J., et al. (2015). Optimized exosome isolation protocol for cell culture supernatant and human plasma.Journal of Extracellular Vesicles, 4, 27031.
8.Zhang, H., et al. (2018). Identification of distinct nanoparticles and subsets of extracellular vesicles by asymmetric flow field-flow fractionation. Nature Cell Biology, 20, 332–343.

La mejor actitud que podemos adoptar es la de trat...

The research team observed changes in head circumf...
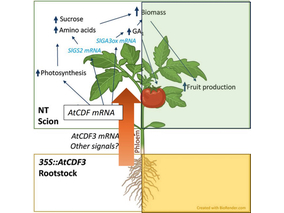
AtCDF3 gene induced greater production of sugars a...
En nuestro post hablamos sobre este interesante tipo de célula del...

Telum Therapeutics, a biotechnology company leveraging proprietary met...